Antecedentes (metodologia ABP):
1910
Flexner presenta su:
Reporte sobre la educación medica
En el cual se propone que se enseñe de la siguiente forma:
/La enseñanza de los procesos biológicos, dentro de las asignaturas bascas *teoría*
/La enseñanza de las alteraciones de los procesos biológicos, en las asignaturas clínicas (practica)
60'
Universidad Mc Master, Canadá.
Universidad Case Western Reserve, USA
Como propósito:
es evitar la confrontación entre la forma de aprender durante los estudios y la forma de trabajar en la vida real.
Metacognicion:
Razonamiento de alto nivel que implica un control activo sobre los procesos cognitivos implicados en el aprendizaje.
Accion de pensar sobre el propio pensamiento.
Metodología abp-utilidad:
Combinar y sintetizar la información presentada en la situación en una o mas hipótesis explicativas; estas hipótesis
no son diagnósticas sino que tratan de dar explicar la información presentada en la situación o problema.
Identificar necesidades de aprendizaje relacionadas al conocimiento, a las habilidades y a las actitudes.
A partir de lo aprendido, identificar los principios y conceptos para poder aplicarlos a otras situaciones o problemas.
M.A. estrategias y habilidades a desarrollar (ABP):
Habilidades a desarrollar:
Comunicar los resultados de una investigación en forma oral, gráfica y por escrito.
Razonar critica y creativa mente.
Tomar decisiones razonadas y analizar en situaciones originales.
Identificar, encontrar y analizar la información requerida para una tarea particular.
Comunicar ideas y conceptos a otras personas.
Lograr la auto confianza necesaria para usar sus habilidades de comunicación y pensamiento en un grupo de personas.
Sustento teórico:
El entender de la realidad surge del medio ambiente.
El conflicto cognitivo estimula el aprendizaje.
Comparación y relación:
Características esenciales de un problema:
Comprometer interés.
Fomentar toma de decisiones , juicios e información lógica y fundamentada.
Cooperación de todos los integrantes.
Ligar aprendizaje previo.
Incorporar objetivos del curso.
Evaluación:
Se fundamenta el constructivismo, la evaluación debe estar
pendiente de todo proceso de construcción del conocimiento y no solo del final.
Debe cubrir:
Los resultados del aprendizaje de los contenidos programados.
El conocimiento que el alumno aporta al proceso de razonamiento del equipo.
Las interacciones personales del alumno con sus compañeros.
Las actitudes frente al proceso de aprendizaje.
La toma de decisiones como actitud de conducción en el aprendizaje.
Hipótesis
Explicaciones fundamentadas con base en la
explicación proporcionada por el caso a través de las pistas y problemas planteados según el nivel de conocimientos.














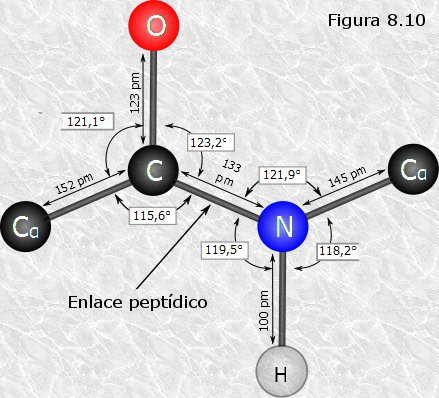




 Los péptidos sólo podrán cambiar de conformación mediante el giro en torno a los enlaces sencillos en los que intervienen los Catetraédricos (Figura de la derecha).
Los péptidos sólo podrán cambiar de conformación mediante el giro en torno a los enlaces sencillos en los que intervienen los Catetraédricos (Figura de la derecha). Cuando se representan los valores de phi (f) y psi (y) de cada AA se obtiene la gráfica de Ramachandran (Figura de la izquierda), en la que se distinguen tres regiones con valores "permitidos" de los ángulos de torsión ya que no dan lugar a impedimentos estéricos en la estructura.
Cuando se representan los valores de phi (f) y psi (y) de cada AA se obtiene la gráfica de Ramachandran (Figura de la izquierda), en la que se distinguen tres regiones con valores "permitidos" de los ángulos de torsión ya que no dan lugar a impedimentos estéricos en la estructura.
